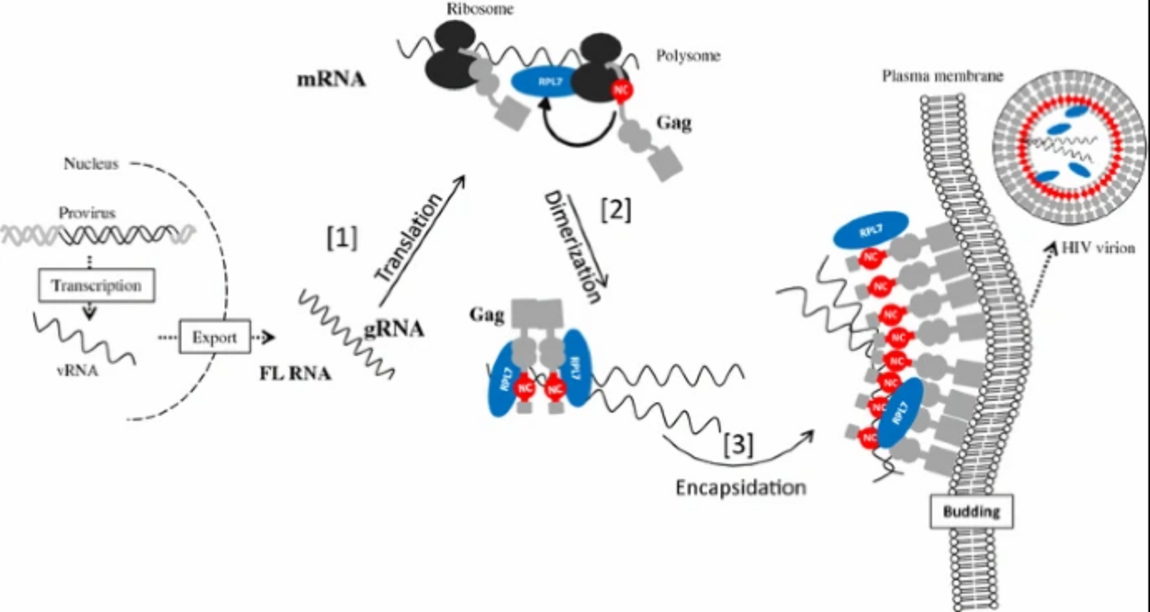
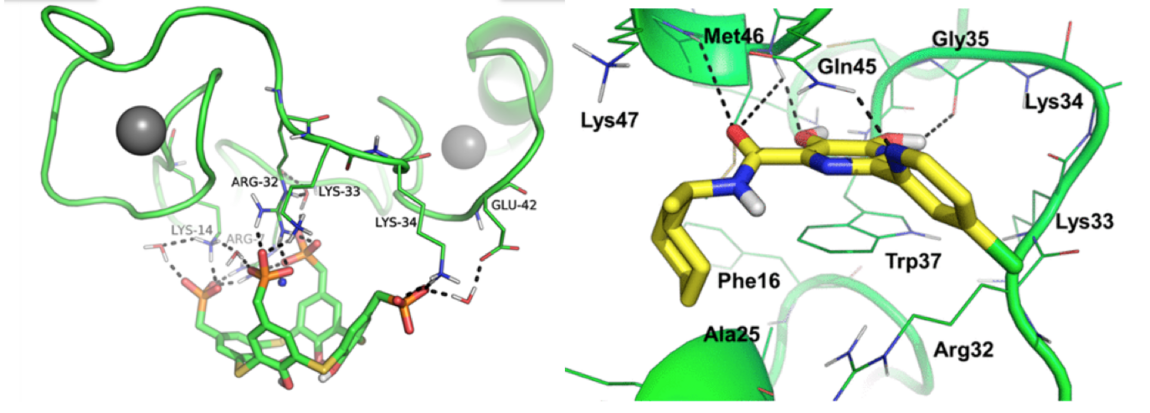
1. Intracellular distribution and roles of NCp7 during the early steps of the retroviral cycle
The nucleocapsid protein NCp7 plays key roles in the condensation and protection of the genomic RNA (gRNA) and the viral DNA. NCp7 also chaperones the reverse transcription and integration steps during HIV-1 replication (for a review see 1,2). To further understand the properties and functions of NCp7, we have explored several research directions.
1.1. Role of NCp7 in viral particles
Using the recently developed 2-thienyl-3-hydroxychromone (3HCnt) and thienoguanosine (thG) nucleoside surrogates, we investigated in depth the NCp7-promoted (+)/(-)PBS annealing of the second strand transfer reaction in reverse transcription (3). These fluorescent nucleoside surrogates were introduced in the (-)PBS loop. This approach allowed us to recover the full set of kinetic rate constants governing the (+)/(-)PBS annealing both in the absence and the presence of wild-type and mutant NCp7 proteins. Through a specific binding, NCp7 exposes the PBS nucleotides to the solvent, allowing the formation of a loop–loop kissing complex. NCp7 also destabilizes the stem, which facilitates the conversion of the loop–loop intermediate into the final duplex.
In collaboration with F. Westerlund (Gothenburg), nanofluidic channels were used to investigate at the single molecule level the delicate balance between DNA condensation and chaperone activity of NCp7 (4). The first ten amino acids were found to be important for both activities. Moreover, we also investigated the initial step of compaction of single dsDNA molecules (Jiang et al, submitted). NCp7 was found to initiate compaction from the ends of long dsDNA, but compaction can also occur in the interior of the dsDNA, preferentially at its most flexible AT-rich regions.
1.2 Cytoplasmic Release of NCp7 from viral particles
To visualize viruses in the host cell during infection, we developed a specific labeling protocol for correlative light-electron microscopy (CLEM). This aim was quite challenging because the labelling approaches developed for fluorescence microscopy are usually not suited for transmission electron microscopy (TEM). To overcome this bottleneck, we adapted a labeling approach with a tetracystein tag (TC) and a biarsenical resorufin-based label (ReAsH) for monitoring the HIV-1 particles during the early steps of HIV-1 infection by CLEM (5). In this approach, the ReAsH fluorophore triggers the photo-conversion of 3,3-diaminobenzidine tetrahydrochloride (DAB), generating a precipitate sensitive to osmium tetroxide staining that can be visualized by TEM. HeLa cells, infected by ReAsH-labeled pseudoviruses containing NCp7-TC proteins exhibit strong fluorescent cytoplasmic spots that overlap with dark precipitates in the TEM sections. Using this approach, single viral cores were observed all over the cytoplasm, and notably near microtubules and nuclear pores.
Using the same HIV-1 pseudoviruses containing NCp7-TC proteins labeled by FlAsH, we quantitatively monitored the NCp7 concentration in the viral cores during the early stages of infection 6. This approach was based on the strong quenching of FLAsH resulting from the high packaging of NCp7-TC in the viral cores. A strong fluorescence increase of individual FlAsH-labeled pseudoviruses containing NCp7-TC proteins was observed in infected cells at 8 and 16 h post-infection. This increase was abolished by inhibition of reverse transcription indicating that the release of NCp7 molecules from the viral complexes is likely connected to viral DNA synthesis. NCp7-TC release was more pronounced in the perinuclear space, where capsid disassembly can occur.